3) 魚類の性分化および性転換機構
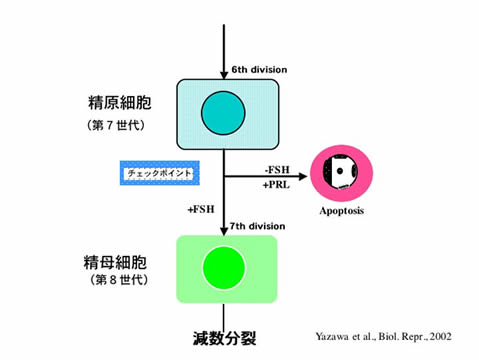
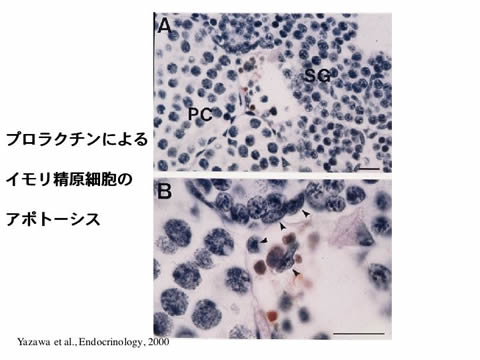
ヒラメの性は基本的にXX(♀)とXY(♂)によって決定されます。しかしながら、性分化時期の温度や性ホルモン処理により容易に性転換を誘導することができます。したがって、ヒラメは、性分化及び性転換機構を調べる上で、大変有効な実験動物です。性分化における性ホルモンの役割を明らかにするため、ヒラメからアロマターゼ(アンドロゲンをエストロゲンに変換する酵素)、エストロゲンレセプターaとb、11b-水酸化酵素(魚類特異的アンドロゲンである11-ケトテストステロンの合成に関与する酵素)の遺伝子を単離し、温度コントロールにより誘導した雌雄の生殖腺におけるこれら遺伝子の発現を調べました。その結果、アロマターゼ遺伝子の発現は、性分化開始以降、雌では急激に増加しましたが、雄では減少しました。一方、11b-水酸化酵素遺伝子の発現は、雄においてのみ、性分化後に増加しました。さらに、性分化時期の遺伝的雌にアロマターゼ阻害剤または抗エストロゲン剤を投与することで生理的雄へと性転換したことから、雌への性分化にはエストロゲンが不可欠で、雄への性分化にはエストロゲン合成の抑制が重要であると考えられました。現在、アロマターゼ遺伝子の発現制御機構の解析及び雄への分化誘導因子の探索を行うと同時に、メダカを用いた機能解析システムの開発にも着手したところです。
Mechanism of sex differentiation and sex reversal in fish
The genetic sex determination mechanism in the Japanese flounder (Paralichthys olivaceus) is basically XX(female)-XY(male) type. The genetic females can be sex-reversed to phenotypic males by rearing the larvae at high or low water temperature or by treatment with sex steroid hormones during the sex differentiation period. Therefore, the flounder provides an excellent model to elucidate the mechanism of sex differentiation and sex reversal.
To address the roles of sex hormones in gonadal sex differentiation, we isolated cytochrome P450 aromatase (P450arom, an enzyme that catalyzes the conversion of androgens to estrogens), estrogen receptor-_ (ER__, ER_, and 11_-hydroxylase (P45011_, the first enzyme involved in fish-specific androgen synthesis such as 11-ketotestosterone) cDNAs and examined their expression patterns during gonadal sex differentiation period. After the initiation of sex differentiation, the level of P450arom mRNA increased rapidly in the female gonad, whereas it decreased slowly in the male gonad. In contrast, P45011_ mRNA level increased only in the male gonad after the sex differentiation. Moreover, treatment of an aromatase inhibitor or an ER antagonist induced the masculinization of the genetic females. These results indicate that estrogen is indispensable for ovarian differentiation, whereas diminution of estrogen biosynthesis is important for testicular differentiation. We are currently studying on the regulatory mechanism of P450arom gene expression and searching inducer(s) for testicular differentiation. |