purescience
第12号 2017年11月
「植物の葉緑体の進化と細菌の細胞壁ペプチドグリカン」
約38億年前に地球に最初の生命が誕生した後、27億年前までに光合成を行うシアノバクテリア(藍藻)が、さらに約10億年前までに、真核生物の一種がシアノバクテリアを細胞内共生させた植物の祖先が誕生し、後にこのシアノバクテリアが「葉緑体」に進化したと考えられています。この祖先が現在の地球上に存在する全ての植物の祖先です。ですが、現在の植物の葉緑体で働くタンパク質をコードする遺伝子の一部をよく調べてみると、その由来はシアノバクテリアだけではなく、別の細菌由来と思われるものが多く存在していることが示唆されており、葉緑体の進化の過程で様々な細菌の共生により遺伝子の水平伝搬が起きた可能性が指摘されています。葉緑体は二重の包膜をもちますが、これは宿主細胞に細菌が取り込まれることにより、宿主の細胞膜が共生体の外側の包膜となり、元々のシアノバクテリアの細胞膜が共生体の内側の包膜となったことによります。一方、全ての細菌にはペプチドグリカン層(PG層)と呼ばれる細胞壁があります。これには細菌を外界の物理的ストレスから防御する役割や、細菌の形を決める機能があります。では、細胞内共生をしたシアノバクテリアの細胞壁はどうなったのでしょうか?細胞内共生により誕生した祖先的な植物は、その後、灰色(かいしょく)植物、紅色植物、緑色植物と3つの植物の系統に進化していきました。このうち、灰色植物のシアネレと呼ばれる葉緑体には、共生体起源のPG層が残っていることが知られていましたが、紅色植物、陸上植物では、進化の初期に失われたと考えられていました。
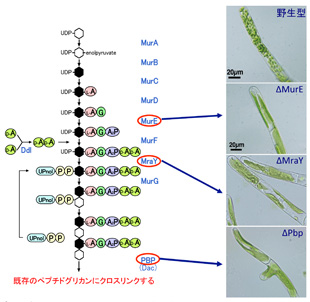
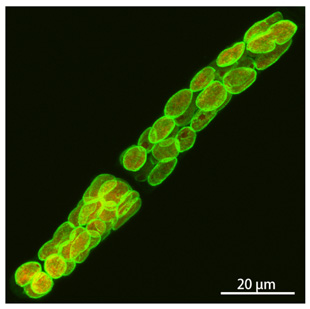
高野教授と私のグループは、この共生体起源のPG層を合成するのに必要な遺伝子群が一式、コケ植物蘚類ヒメツリガネゴケに存在し、葉緑体で機能していることを発見しました。またこの遺伝子一式は、陸上植物の基部に位置する緑藻類の中でも、陸上植物の祖先となったグループにしか見られません。陸上植物ではシダ植物の一部や、裸子植物にもこの遺伝子群が残っていましたが、被子植物には一部の遺伝子しか残っていませんでした。では、植物の葉緑体にPG層は存在するのでしょうか?コケの葉緑体を、電子顕微鏡で観察したところ細菌のPG層のような厚い構造は見当たりません。PG合成を行うための遺伝子群は残っていても、PG層を作っていないのでしょうか?ヒメツリガネゴケは相同組換えという技術を使って自由に目的の遺伝子を改変することができるので、PG合成系の遺伝子のいくつかの機能を失わせてみたところ、葉緑体の分裂が阻害されて、巨大な葉緑体が出現することが分かりました(図1)。すなわち、ヒメツリガネゴケではPG合成系の遺伝子は葉緑体分裂に関与することがわかりました。この機能は細菌でも同様で、PG合成系を阻害する薬剤を処理すると分裂が阻害されてしまいます。やはりこれらの遺伝子はコケの葉緑体の中でもPGを作っていそうです。そこで我々はPGの前駆体を化学的に修飾し、PGを蛍光標識することにより、葉緑体のPGを可視化することに成功しました(図2)。現在、葉緑体からPGを単離することを目指して、研究を進めています。
一方、先にも述べたように被子植物のゲノムにはPG合成系遺伝子は一部しか残っていません。MurEと呼ばれる合成系遺伝子は全ての陸上植物に残っているのですが、PG合成できないのに、どうして残っているのでしょうか?シロイヌナズナという被子植物のモデル植物を使って調べたところ、シロイヌナズナではMurE遺伝子が壊れると葉緑体が正常に発達せず、アルビノという白い植物体になることが分かりました。コケのように巨大葉緑体は生じません。シロイヌナズナではMurEは、葉緑体ゲノムにコードされた遺伝子の転写に関与することが分かりました。このように遺伝子が元々の役割とは全く異なる役割をもつようになることは、生物進化の過程でよく起きます。遺伝子もリユースできるものは、リユースされているのですね。